We are so excited to announce the agenda and speakers for our 2019 Interdisciplinary Integration Symposium, titled "Airway Oscillation: An Interdisciplinary Approach to the Production of Voice, Airflow, and Resonance". The 11th Annual Interdisciplinary Integration Symposium will be held in Lincoln, NE on April 11-12, 2019.
The speaker line up and two day agenda are outlined below. To learn more about the course (including description and objectives), and to register online, CLICK HERE. Don’t wait to sign up, this one is sure to fill up!
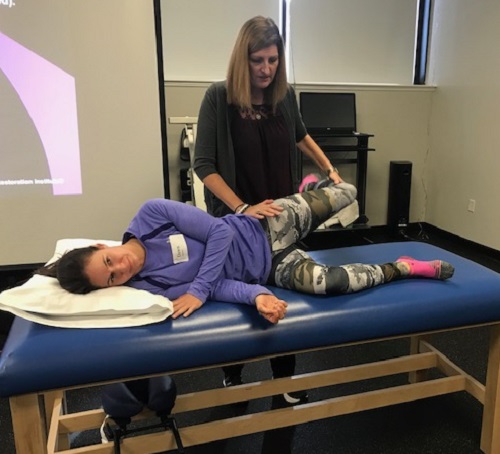
Recently returned from the greater Chicago area for Pelvis Restoration at The Trainers Club where I was welcomed by Wynne Conklin. Fellow PRC and faculty member Dan Houglum served as our lab assistant for the course and my stay at the Houglum Hotel was first class by any measure, as was his presence at the course. Newcomers, many who were mid-stream with introductory courses and a couple savvy vets who’d been to many secondary courses were dialed for a PRI steak of a course.
As is generally the case, the pelvis restoration discussion was one of integrating pelvic and thoracic diaphragms with great discussion about how to implement with athletes of all ages and capacities. Many thanks to Eric Hrycko for his willingness and outright enjoyment of lab demonstrations as well as the many others who contributed during lab demonstration and discussion of PEC and Left AIC underlying neuromechanical patterns that elicit compensations of immensely varying sorts.
We discussed how to best address the "big ones" first in terms of identifying inhibition strategies after identifying potential treatment options via administering PRI special tests. This group was particularly effective and variable in lab, showcasing a broad array of normal pelvis asymmetries that allowed a thorough and comprehensive lab demonstration that is not always possible at every course. Literally the entire group participated in discussions and/or lab on at least one occasion during this course–awesome. Thank you all for being such involved students of this fine science!
Thanks again Dan and Wynne for taking the time and expending the energy to ensure that this was a great environment to take in the science of Pelvis Integration! Thoroughly enjoyed Pelvis Restoration Chi-town!
I liken teaching Myokinematics of the Hip and Pelvis to putting on a comfy pair of slippers. I took my first Myokin course back in 2001 the first time Ron offered this material. The course has evolved over time as the Institute learns more about our normal balanced asymmetry and how we compensate in three planes. Despite our deeper understanding of how the visual, occlusal position and proper shoes affect compensation patterns, clinicians still need to understand the fundamentals of a Left AIC pattern as it relates to position and performance. An F (femur) moves on an A(acetabulum), yes, but an A also can move and stabilize an F. This is the foundation of this course. We then need to fully understand how our patients compensate for this normal pattern. Did they or did they not over lengthen their anterior hip ligaments? For me, this is the question of the 2 day course!
In Worcester Mass this weekend, I had an amazing group of clinicians. A large number were taking their first official PRI class. So Fun! But, all of them knew of PRI or were working along side a co-worker using PRI in their practice. They were all in Worcester to learn for themselves how to help their patients/ athletes.
Once we were all clear on the pattern and the position of AF whether it be in IR or ER, we learned the tests to determine their positions and compensations. On Sunday afternoon, we just problem solved our way through the treatment progressions. The class was focused on utilizing functional tests and muscle algorithms to guide what came next. For years I used an analogy in my teaching that seeing a patient present in the clinic as a L AIC was like seeing Yellow VW bugs driving past you on the road. But it is a new day and time for a new analogy, that I now refer to as "Mustang Sally". We had some fun after lunch taking our group shot with my Mustang rental car that I’ve been upgraded to, 2 weekends in a row! New car for my husband Chris, I think maybe! Since then, I see Mustang’s everywhere I go, now that I am looking for them!
Hats off to the staff of Central Mass Physical Therapy! This is the third time I have taught at this host site. I was able to meet some new staff, make some new friends and see some familiar faces. Damion Perry, Yolanda Pappas, Borbala Suranyi and Gene Lyons were all in my Pelvis Class last month in Boston. Meredith Stephens, MS, PT is also a faculty member for Anatomy Trains. We had some great gait discussions at lunch. I look forward to learning more from her in the future. Maureen Watkins, PT is faculty at Northeastern University in the PT department. Her non-patho hip helped me demonstrate how to develop hole control on the right side with a glut max and obturator in the transverse plane. I always enjoy teaching Myokinematics. That’s a wrap for 2018. I will look forward to teaching this awesome course next year!
The need to position the left hemi diaphragm was introduced right off the bat to ensure that the right diaphragm autocracy stops ruling our autonomics! Other key concepts included the need to facilitate hamstrings and IOs/TAs to establish a left ZOA followed by reach activities to hold the ZOA and direct air into previously-restricted areas of the rib cage. A series of gait videos enabled us to view firsthand various pelvic and thorax movement strategies. Lack of arm swing on one or both sides were reflected in our PRI tests. They served as a reminder that arm swing is 50% of the gait cycle!
Thank you to Kathryn Bragg, PT, and Laurie Johnson, PT, seasoned clinicians tuned into nuances of performance of PRI non-manual techniques. The take-home from that discussion: So long as the reference centers are sensed by the patient, stated in the instructions as “You should feel…”, the exercise can be quite effective. Ultimately fine-tuning areas that are holding tension and tweaking alignment can help the process.
With Jill Tender, DPT, as our superior T4 model, we succeeded in unearthing a compensatory respiration strategy that responds to additional non-manual and manual techniques beyond what is needed to manage those who have not yet resorted to this compensation. Others who enriched our learning-through-demonstration were: Amelia Franklin, DPT, OCS; Benjamin Fuentes, PT; Tara Pickett, DPT, GCS; Jacqueline Richards, DPT; and Karthik Yadagiri, PT. I really enjoyed my weekend with this wonderful group of clinicians. They gained an appreciation for just how influential alternating, reciprocal rib cage movement is to efficient movement and to the overall health of human physiological systems….and collectively rejoiced in the resolution of wallet-gate.
Ron Hruska and I attended the 2018 International Association of Orofacial Myologists (IAOM) this weekend in Charlotte, NC. It was an informative conference attended by Speech Language Pathologists (SLP), Registered Dental Hygienists (RDH), Dentists and one other PT. The speakers included physicians (MD and DO), dentists, and SLP’s who spoke about topics including but not limited to: connecting the mouth to the rest of the body, guiding orofacial development in children, techniques for speaking and feeding in children with special needs, techniques for addressing tethered oral tissue (TOT; tongue and lip ties), wound management from TOT surgery, the tongues effect on the body, genetic disorders, and publication trends and evidence in Orofacial Myofunctional Therapy literature.
Ron spoke on Orofacial Myologic Disorders (OMD) Relationship to Postural Asymmetry. It was a well – received lecture where Ron focused on respecting the neck, asymmetry, anatomy, forward head posture and the importance of recognizing sense. He performed a test/ retest demonstration of how a PRI technique results in neutrality. They were amazed to say the least!
Following this lecture Ron and I received many questions from the course attendees about the value of physical therapy. We tried hard to explain to them that their local PT without PRI experience may not be able to address their patient’s issues as demonstrated by Ron in the lecture. It is likely these course attendees (SLP’s and RDH’s) will be contacting you from the PRI website as a clinician to help them treat their patients. Understand that they believe they are the ones who should be treating the tongue and mouth directly. It will be a nice opportunity for PRC’s to help guide these practitioner’s understanding how the position of the neck and body, and the underlying asymmetry, we all appreciate, can have a significant influence on their treatment, approach and results when addressing oral myology pathology and disorders. We both came away with a high degree of respect and appreciation for these interactive and collaborative, multi-disciplinarians.
The title of this artistic illustration reflects how our two lateralized hemi-rib cages operate when their top column of ribs move away from the central axis, or the spine. Bifarious is a botanical name for a structure that has parts arranged in two rows on either side of a central axis. The Latin definition of ‘bifarious’ is ‘double’. These two representations of the anterior chest wall help clarify the likely degree of lateral flexion or thoracic wall abduction of each row of ribs, with respect to the lung, organ and neurologic asymmetry guidance. It reminds the viewer that they are not looking at one rib cage when you look at the chest or thorax. They are seeing double. Two chest walls that revolve and rotate around a central axis, making all our chest walls bifariously oriented.
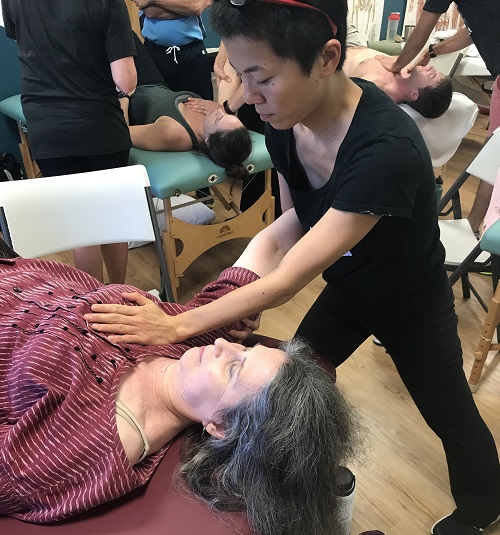
The definition of -ation is something connected with an action or process, or the process of doing something. I feel that I am doing this every time I teach this course. Processing references with desirable action to achieve outcomes that can become reprocessed through the integration of sense of this -ation. This can become a revelation for those who are struggling to change processing. Once again I appreciate Finish Line Physical Therapy for hosting another course in NYC. Tying the last hour of the course, humeral – scapular – thoracic activity with the first hour of the course, calcaneus activity allows the attendees to complete a full circle of sensory referencing, clinically and personally. This group was great in not only participation but in helping Kentaro Ishii MS, ATC/L, PES, CE, PRT rethinking about course organization. Michael Macchiarulo PT, DPT and Monica Saenz DPT thank you for representing the RAIC/LBC and LAIC/RBC pattern references throughout both days.
Monica Integrating R AIC/L BC references while Michael integrates L AIC/R BC references.
Michael integrating R AIC/L BC references while Monica integrating L AIC/R BC references.
Brynn Fessette DPT, FAFS, Joshua DiLoreto ATC, and Chris Leib DPT, CSCS,Cert MD were awesome lab demonstrators and volunteers and enriched the course by their willingness to share their ‘sense’ of what they were feeling and integrating.
But the best thing about teaching, is meeting new people, hearing new reasoning, and integrating new concepts based upon novice and novel experience. Boris Dugandzie and Dino Dogan were two people who exposed me to a culture and country that I knew vey little about. Because of them, Croatia is now on my travel destination bucket list. Their attendance helped me find a different kind of pronation and supination between two Croatians, who have little need to reach forwards or upwards with their arms and hands. Ron demonstrates the -ation, between the floor and the thorax between two Croatians, who struggle to find it.
It is always comforting to be teaching PRI regardless of location, but last weekend I was in Detroit, MI. My wife’s family are all from just west and south of the Detroit area, so I was in very familiar surroundings. And the host group of Team Rehabilitation was an excellent bunch to host Myokinematic Restoration.
With a full room of nearly 40 health care professionals and movement specialists, and nearly all in attendance having never heard PRI previously, it was an awesome learning environment for everyone. This group was very dynamic and asked great questions. As this course evolves, we end up adding more and more lab time. This past weekend, 1/3 of the course was lab based, so we had ample opportunity to learn from each other.
We started our dive into the difference between normal pelvifemoral mechanics around the normal neuromechanical presentation that is the L AIC pattern. Then we went deeper into normal compensation vs. pathological compensation patterns. We transitioned into muscular ramifications of the inability to get out of the L AIC pattern, which highlighted the importance of having a "good boy band" rather than a "bad boy band."
We had ample time to practice the positional testing as we were able to determine what patterns we all had inside of us. Low and behold, every single one of the attendees proved to be in a PEC pattern. Based on this group of attendees all having the same extended pattern, we had to have the "Batman vs. Bruce Wayne" conversation. I was in a room of "Batmans”, and we all needed to be more like "Bruce Wayne." This necessitated a demonstration of how to get someone out of a PEC pattern and into a L AIC pattern, so the rest of the weekend had an opportunity to resonate with the attendees. With two activities, we were able to take a very strong PEC individual, and turn them into a L AIC pattern. Because underneath the bilateral extended patterns of the PEC lies a L AIC pattern.
The "PEC busting" demonstration helped us proceed into the management of a L AIC pattern. Nearly the entire afternoon on Sunday was lab time, where everyone had the opportunity to find and feel hamstring, IC Adductor, gluteus medius, internal obliques, and gluteus maximus activity. Reports of "feeling different" and "feeling my hips" were common comments after our lab.
My thanks to Team Rehabilitation for their hospitality. They know how to host a course! My thanks to Shelly DeRuiter and Craig Stasio, who were awesome lab assistants. Thank you to Cyril Shuster, Alexandre Vieria, Todd Cummings, and Michele Weis for your excellent questions and re-states. Thank you to Michelle Shrader and David Selak for allowing us to learn from you in our demonstration portions of the class. My hat is off to Alexandre Vieria, as he traveled from Brazil to hear PRI! I greatly appreciate our conversation around tennis players and the necessity to have excellent frontal plane integration. Thanks to everyone for great weekend!
Thank you Ryan Murray and the rest of the Fortius Athletic Staff for hosting Pelvis Restoration this past weekend. Also, a huge thank you to Jon Rowe, CSCS, PRT, for being my lab assistant.
This class was awesome with their questions, desire to learn and their attention the entire weekend. This course is challenging from the sheer amount of information given and you guys did great. Pelvis Restoration goes into great detail with position, muscle inhibition/facilitation and respiratory concepts behind "AFIR" and "AFER." I feel the concept of push/pull and asymmetry between the left and right side for alternation during the gait cycle hit home. Vancouver is a beautiful city and I hope to come back and visit. Thanks again!!
It’s not too late, but the Postural Restoration Trained™ (PRT) credentialing program application deadline is quickly approaching! If you are interested in applying this year, applications are due on October 15th and testing will take place on January 7-8, 2019 in Lincoln. To learn more about eligibility for PRT credentialing and download the application, CLICK HERE!
What have others shared about their experience with PRT credentialing?
"The PRT credentialing process was a wonderfully fulfilling experience. The challenge of putting together the application combined with the nurturing environment of the testing phase in Lincoln was such an impactful learning opportunity. All of the Institute’s staff members involved with the process in addition to my fellow applicants were fantastic in both professional quality and character. I am certainly proud to have earned the PRT designation in the eyes of the Postural Restoration Institute." – Ryan DiPanfilo, ATC, CSCS, PRT
"Obtaining the PRT was more than just taking another test to add letters behind my name. It was an opportunity to challenge myself to move to the next level in an area I am passionate about. The 2 days at spent at PRT testing weren’t the end of over 2 years of studying, but the beginning of moving forward with an organization that is quietly changing the way we treat the human body. I highly encourage anyone who has met the PRT requirements to take the next step to obtain the PRT credentials." – Tim Dempsey, CSCS, PRT
"Everything was great! Really good stimulated critical thinking and application of the information. The two days flowed really well and built off of everything before it. The application process was a great learning experience in itself." – Andrew Hauser, ATC, CSCS, PRT
I enjoyed teaching Myokinematics of the Hip and Pelvis last weekend in Asheville, NC! This was a special weekend as I was able to take Mr. Jen Poulin aka Chris with me to help! It has been awhile since we were able to travel and teach together. We were joined by Josh Owen, ATC, PRT. It was great to meet Josh and have him help with the labs.
This course was hosted by the fun energetic group of clinicians from the Charles George VA Hospital. Tammy, Doug, Amy, Monica to name just a few of the staff, made us feel welcome.
Most of the class were taking their first PRI course! I always enjoy this class as it really hits home the fundamental concepts of AF and FA position and common compensations. We all appreciate the importance of respiration as it relates to stabilization of a pattern. This course really helps the course participant understand how respiration and gait are so interconnected. Concepts of swing vs. stance, AFIR, AFER and soft tissue compensations were understood as it relates to position, pattern and performance. I had so much fun teaching this past weekend! I can’t wait to come back!
I will get a few weeks off and look forward to heading up to Worcester, Mass on Oct. 13-14th to teach this awesome course again!
Thanks for everyone’s prayers for NC. Hurricane Florence left her mark, but we are Carolina Strong!
Regardless if you are a patient, a course attendee, a guest lecturer, or going there for consultation, when you enter the doors of The Cantrell Center for Physical Therapy, Sports Medicine and Wellness, you are family. The entire staff were engaging and enlightening. All of the course attendees accommodated my needs to embed the center of our orientation, the orientation for autonomics, and the conduit of zones, i.e. the foramen magnum and the vertebral foramen of the atlas, in organized tri-planar revolving movement. They helped me teach future attendees that will appreciate the needs to embrace laterality, asymmetry and sensory integration for cervical function. The content of this course reflected the attendees willingness to consider the reasoning for the neck to revolve, as well as the reasoning to restore appropriate universal tension provided or placed on the neck and its contents. There was a wonderful example of a left Side Bend positioned cranium in class that was so willing to allow me to demonstrate re-positioning and re-patterning techniques with it. Courtney Stearn DPT owns this cranium and she and her cranium, cervical spine and cortex were anatomical gifts to all of us. Her personality and behavior made the first day a complete day of enjoyment and learning. Thank you Courtney! Sitting to the right of Courtney was Amy Brown DPT who owned a cranium and neck that reflected right torsion through the sphenoid. She also contributed so much to the second day by allowing me to discuss her testing outcomes and her technique demonstration. She could not have been in a better place or in a better time for this presenter. Thank you Amy, we are all grateful for your attendance and willingness to share your head and neck.
Course participation, by everyone, and I mean everyone, was awesome. So thank you for making this weekend a very special one for me. I can still see and hear Tassie Cantrell PT putting out strawberries and blueberries, putting furniture away afterwards, smiling ALL the time and answering questions on when she and her facility will be putting on another PRI course, and asking questions about O on A and A on O.
Lola Rosenbaum DPT thank you for being my “sphenoid” and Leah Whipple DPT thank you for being my “shift”. And finally, Skip George DC, PRC, thanks for being there, and mentoring me, and guiding me, and listening to me, and assisting me, and putting up with my metaphoric satire, and for loving me.